
Introdução
A visão é responsável por cerca de 75% de nossa percepção. Resumindo de
forma extremamente sintética o ato de ver é o resultado de três ações distintas: operações óticas, químicas e nervosas.
O órgão responsável pela captação da informação luminosa/visual e
transformá-la em impulsos a serem decodificados pelo sistema nervoso é o OLHO:
é um instrumento altamente especializado e delicadamente coordenado, e cada
uma de suas estruturas desempenha um papel específico na transformação da
luz, se transformando no sentido da visão. Toda a entrada de luz do meio externo
até chegar à retina, faz parte do sistema ótico, propriamente dito. A
sensibilização da retina se faz quimicamente, a luz convertida em impulsos
elétricos, é transportada através do nervo ótico até o córtex.
Anatomia do Olho Humano
O olho humano é formado por um conjunto complexo de elementos que atuam
de forma específica para que o ato de olhar, ver ou enxergar ocorra.
Primeiramente existem aquelas estruturas responsáveis pela captação da luz e
desempenham função ótica, posteriormente aparecem os elementos que
transformam o impulso luminoso em impulso elétrico, através de reações
químicas. De forma simplificada o olho é formado por: córnea, íris, pupila,
cristalino, retina, esclera e nervo ótico.
Córnea: É a primeira estrutura do olho que a luz atinge. A córnea se constituide cinco camadas de tecido transparente e resistente. A camada mais externa, o
Epitélio, possui uma capacidade regenerativa muito grande e se recupera
rapidamente de lesões superficiais. As quatro camadas seguintes, mais internas,
são que proporcionam uma rigidez e protegem o olho de infecções.
Íris: A porção visível e colorida do olho, logo atrás da córnea. Possui músculos
em disposição tal que possam aumentar ou diminuir a pupila, a fim de que o olho
possa receber mais ou menos luz, conforme as condições de luminosidade do
ambiente.
Pupila: É a abertura central da íris, através da qual a luz passa para alcançar
o cristalino.
Cristalino: É quem ajusta na retina o foco da luz que vem através da pupila.
Tem a capacidade de, discretamente, aumentar ou diminuir sua superfície curva
anterior, a fim de se ajustar às diferentes necessidades de focalização das
imagens, próximas ou distantes. Esta capacidade se chama "acomodação".
Retina: É a membrana que preenche a parede interna em volta do olho, que
recebe a luz focalizada pelo cristalino. Contém fotorreceptores que transformam a
luz em impulsos elétricos, que o cérebro pode interpretar como imagens. Existem
na retina dois tipos de receptores: bastonetes(+ ou -120 milhões) e cones (+ ou - 7
milhões), que se localizam em torno da fóvea. Cada receptor comporta em torno
de 4 milhões de moléculas, ricas em rodopsina, que é capaz de absorver quanta
luminosos decompondo-se em duas outras moléculas.
Nervo Óptico: Transporta os impulsos elétricos do olho para o centro de
processamento do cérebro, para a devida interpretação.
Esclera: É o nome da capa externa, fibrosa, branca e rígida que envolve o olho,
e contínua com a córnea. É a estrutura que dá forma ao globo ocular.
A Fisiologia do Desenvolvimento da Visão
A visão é feita pelo cérebro. Os olhos funcionam como órgãos de conversão
seletiva do estímulo luminoso em sinais elétricos. Durante todo o trajeto através
do sistema visual, os estímulos vão sendo depurados até gerarem uma impressão
visual única, provavelmente no córtex occipital. Existe um período da vida em que
esse processo se desenvolve e no fim do qual se consolida, chamado Período de
Maturação Visual. Didaticamente, dividimos a visão em central e periférica.
A visão central da criança, do nascimento até cerca de oito anos de idade,
comporta-se diferentemente da do adulto: ela aperfeiçoa-se ou deteriora-se com a
qualidade da informação visual. Nessa fase, conhecida como “período dematuração”, o cérebro interage abertamente com a retina para melhorar a
interpretação das informações do ambiente. É fundamental, pois, que ele receba
informações claras e precisas nesse período.
No entanto, isso só é possível se ambas as retinas transmitirem sinais nítidos e
semelhantes.
Como cada olho oferece imagem de um ângulo diferente, o cérebro acaba
recebendo duas imagens discretamente díspares. Quando as une numa impressão
visual única, a disparidade gera um efeito tridimensional. Esse fenômeno só é
possível em virtude da mistura de informações das duas retinas, promovidas pela
fibras dos nervos ópticos. Quando isto não ocorre, como em casos de estrabismo, o
desalinho dos eixos visuais faz com que cada olho forneça imagens muito
diferentes entre si, conflitantes, impedindo o processo de fusão, o que faz com que
o cérebro acabe "escolhendo" uma das imagens, desprezando a outra. Com isso o
olho que tem sua imagem preterida, não se desenvolve na mesma proporção que o
outro, pois não é exigido, sendo pouco usado. A correção tardia, só beneficia a
estética, pois o período de desenvolvimento da visão já terminou. É dado o nome
de ambliopia à baixa visão decorre de uma deficiência de maturação visual, e ao
indivíduo que a porta, amblíope.
A luz, proveniente de um objeto de interesse, atravessa os meios transparentes
do olho e chega à retina. Aí, ela é convertida em impulsos elétricos, que são
levados ao córtex occipital através dos nervos e vias ópticas. No córtex, os
impulsos são decodificados na forma de uma impressão visual.
A retina não tem a mesma sensibilidade em toda sua extensão. Possui uma
área, do tamanho da cabeça de um alfinete, responsável pela discriminação dos
objetos. Essa área é conhecida como fóvea. A fóvea fica próxima do disco óptico,mas ligeiramente deslocada para o lado temporal. O disco óptico é o local onde o
nervo óptico penetra no olho. Como nessa região não existem fotorreceptores, ele é
completamente cego.
Todo o resto da retina é responsável pela visão de campo. A visão de campo é
fundamental para a locomoção, pois dá uma apreciação de conjunto.
A medida da visão foveal chama-se acuidade visual. A da visão de campo
chama-se campimetria. É importante o conceito de que a perda de visão de campo
é mais desabilitante que a da visão central.
A Fisiologia da Visão Binocular
Binocularidade, em seu sentido mais amplo, é o termo que se aplica à
capacitação de apreender estímulos visuais com dois olhos. Entre as diferentes
espécies que possuem esse atributo, os modelos da relação binocular, suas
peculiaridades e fins são, entretanto, muito distintos. Enquanto para coelhos a
função binocular é a de provimento de campos visuais independentes, garantidos
por órgãos visuais de cada lado da cabeça, nos primatas superiores (e no homem),
a percepção visual do espaço se dá com base na frontalização dos olhos. Assim,
coelhos gozam de uma extensa visão de “campo”, cobrindo 360º, um escrutínio
completo do espaço ao redor de seu corpo, o que lhes propicia meios de fuga de
predadores. Já entre nós predomina a superposição praticamente completa dos
campos visuais. Tal superposição, ocasionando perda de 180º da discriminação
visual do espaço relativamente à do coelho traria como vantagem, em
contrapartida, a capacidade de percepção de “profundidades” de objetos nessecampo visual, isto é, a de percepção das localizações egocêntricas desses objetos à
distância deles ao agente da percepção.
A visão binocular de seres humanos resulta da superposição quase completa
dos campos visuais de cada olho, o que suscita discriminação perceptual de
localizações espaciais de objetos relativamente ao observador (localização
egocêntrica) bem mais fina (estereopsia), mas isso ocorre em, apenas, uma faixa
muito estreita (o horóptero). Aquém e além dela, acham-se presentes diplopia e
confusão, sendo necessária supressão fisiológica (cortical) para evitá-las.
A percepção de distâncias egocêntricas não é necessariamente vinculada a essa
superposição de campos, podendo ser dada por uma série de “pistas monoculares”
tais como as de:
a) interposição de estímulos (os mais próximos “cobrindo” os contornos e áreas
dos mais distantes);
b) tamanhos relativos das imagens (maiores para os de objetos mais próximos,
menores para os dos mais distantes);
c) contornos e brilhos (mais acentuados com a proximidade, esmaecidos com o
distanciamento);
d) zonas de sombras e iluminação (sugerindo relevos e cavidades);
e) perspectiva aérea (coloração mais azulada para grandes distâncias, pela
interposição de ar entre o observador e os objetos);
f) perspectiva cinemática (pelo observador em movimento: objetos mais
próximos com deslocamentos aparentes mais rápidos).Fisiologia e Percepção
Percepção é o processamento, em etapas sucessivas, da luz que chega aos nossos
olhos. Esta informação está codificada (não no sentido da semiologia) através de
regras de transformação naturais. Essencialmente, as regularidades destes
fenômenos, referem-se a três características da luz: sua intensidade, seu
comprimento de onda, sua distribuição no espaço (e no tempo).
A interpretação de luminosidade de um objeto, já provém de nossa
interpretação da quantidade se luz emitida ou refletida por ele.
A visão de pode ser, de acordo com a luminosidade: fotópica ou escotópica:
Fotópica: modo de visão "normal", quando são iluminados por uma luz diurna.
Aciona basicamente os cones, e por isso, distingue as cores. Acontece na região
central da retina, diretamente ligada ao movimento da pupila, tem sua acuidade
acentuada.
Estocópica: é a visão "noturna", predominante a atividade dos bastonetes,
apr e s enta uma pe r c epção ac romát i ca, de f raca acuidade e acont e c e ,
principalmente diante da baixa luminosidade, na região periférica da retina.
A percepção da cor se dá através do espectro de luz, o arco-íris, sendo a cor,
propriamente dita da luz emitida varia de acordo com o comprimento de onda. A
variação do comprimento de onda é invisível a olho nu, nas regiões externas do
espectro.
Cor = comprimento de on
ACUIDADE VISUAL
Acuidade visual relaciona-se com a capacidade de resolução espacial de dois pontos e depende da densidade dos receptores na retina e do poder de refração do sistema das lentes ópticas. Distâncias na retina são referidas em termo de ângulo visual. Assim dizemos que a capacidade do olho resolver dois pontos está associada com certo número de ângulo visual. Quando são realizados testes visuais com letras e números de diferentes tamanhos num consultório oftalmológico, está se pesquisado justamente como anda a acuidade visual do paciente. O olho (a região da fóvea) possui uma capacidade tão acurada para descriminação que será capaz de discriminar duas linhas afastadas por 0,29mm a uma distancia de 1 metro !
DEFICIÊNCIAS VISUAIS DEVIDO AOS MECANISMOS DE FORMAÇÃO DE IMAGEM NA RETINA
Denominamos olho emenotrópe aquele que focaliza os objetos sem o uso da acomodação. Mas algumas pessoas não conseguem formar uma imagem com nitidez na retina ao observar objetos distantes parecendo-lhes embaçados. Porém, enxergam bem os objetos próximos. Chamamos esse olho de míope e o uso de lentes divergentes resolve o problema. Outras não apresentam problemas com a visão de objetos distantes, mas apresentam dificuldade para enxergar de perto: é o olho hipermétrope e as lentes convergentes resolvem o problema. O astigmatismo é uma condição onde a curvatura da córnea não é uniforme e a imagem na retina causa sensação de turgidez e a correção se faz com lentes cujos meridianos são compensados por uma lente cilíndrica. A catarata é uma condição em que o cristalino perde a sua transparência, podendo levar a cegueira, sendo considerado um processo de degeneração senil. Entretanto, pode também ser causada pela exposição aos raios X, hiperglicemia, dieta carente de alguns aminoácidos, anóxia, etc.
Além das particularidades com relação ao cristalino o formato do globo ocular (tamanho) leva a déficits de focalização da imagem sobre a retina.
MORFOLOGIA DA RETINA
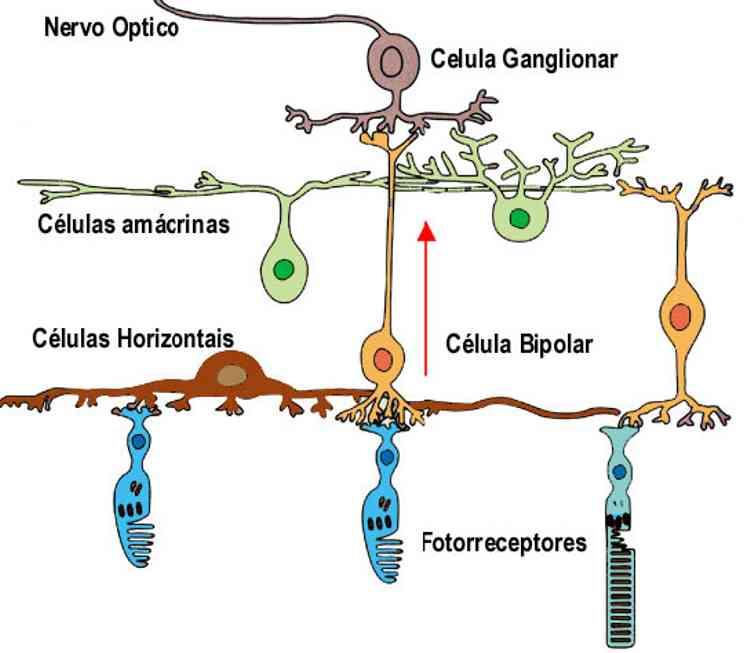
A retina pode ser dividida em uma metade nasal e outra temporal supondo uma linha imaginária passando pela fóvea. Possui varias camadas de células distintas: a mais interna é a camada de neurônios ganglionares cujos axônios formam o nervo óptico (II par craniano). Nas duas camadas nucleares estão os corpos celulares de células especificas: na interna, células bipolares, células horizontais e amácrinas e na externa, fotorreceptores. As duas camadas plexiformes são os sítios de profusa comunicação sináptica. A camada externa é formada pelas estruturas que contem os elementos transdutores dos fotorreceptores.
Repare que a camada de fotorreceptores está voltada contra a chegada da luz. Isto parece paradoxal, mas como as camadas de células nervosas são transparentes, a luz passa por elas com distorção mínima. Além disso, esta disposição evita ao máximo as reflexões luminosas devido à camada pigmentar que absorve totalmente a luz. Já o fluxo de informações se dá no sentido oposto: fotorreceptores ® células bipolares ® células ganglionares. Evidentemente, as células ganglionares conduzem para o cérebro o resultado local e final do processamento visual na forma de PA propagados. As células horizontais também recebem informações dos fotorreceptores e influenciam as células bipolares enquanto as células amácrinas influenciam a excitabilidade das células ganglionares.
FOTORRECEPTORES
Os cones e bastonetes são células sensoriais fotossensíveis e responsáveis pela transdução foto-elétrica. A retina humana possui aproximadamente 120 milhões de bastonetes e 6 milhões de cones Tanto os cones como os bastonetes possuem três segmentos: externo, interno e o terminal sináptico. No segmento externo ocorrem as reações fotoquímicas e no terminal sináptico, a liberação de NT para as células nervosas da retina (células bipolares e horizontais). O segmento externo dos bastonetes possui, em seu interior, uma pilha de discos membranosos (ou lamelas) flutuantes nos quais as reações fotoquímicas ocorrem; os cones possuem segmentos mais curtos, mas o processo é semelhante. Os segmentos externos dos bastonetes e cones estão em constante renovação.
Os bastonetes são sensíveis a um amplo espectro da luz (380 a 650nm); possuem apenas um tipo de molécula fotossensível, a rodopsina que está em concentrações bastante elevadas. Em função dos tipos de pigmentos fotossensíveis a determinados comprimentos de onda há três tipos de cones: azuis, verdes e vermelhos.
Na macula lútea, há uma depressão denominada fóvea onde as imagens são focalizadas com maior nitidez ou acuidade. Essa depressão é devida à presença exclusiva de cones e a luz incide diretamente sobre esses receptores. Esse ponto é arbitrariamente conhecido como o ponto central e constitui a origem da superfície retiniana (0o). À medida que se afasta do ponto central, a densidade de cones diminui drasticamente e o de bastonetes aumenta. Na fóvea a relação de inervação das células ganglionares com os cones é pequena (1:1) e com os bastonetes é muito grande, ocorrendo muita convergência.
O disco óptico é o ponto de emergência do nervo óptico. Isso significa que neste ponto não há retina, portanto uma região “cega”. De fato, possuímos um ponto cego em cada retina. Então como é que não vemos essas falhas em nosso campo visual? É que o córtex visual “preenche” esses escotomas fisiológicos completando a imagem em cada campo visual.
MECANISMO DE TRANSDUÇÃO FOTOQUÍMICA
Bastonetes
Os bastonetes os fotopigmentos denominados rodosina. Ela é sintetizada a partir da opsina (proteina) e o retinal (lipideo) que é derivado da vitamina A. Na ausência da luz, o retineno da rodopsina se encontra na posição 11-cis e na presença de luz, a opsina e o retinal se separam e a opsina se descolore (rosa para amarelo palido). Mais especificamente, a luz tem como função modificar a conformaçaõ da rodopsina 11-cis em 11-trans. A decomposição causa alteração na condutância iônica na membrana do fotorreceptor e provocar um potencial receptor e como consequencia reajustar a liberaçao de NT (glutamato) do fotorreceptor. Como isso acontece?
Quando não há luz incidente nos fotorreceptores, a membrana dos discos tem a sua permeabilidade alta para os ions Na e há uma corrente de despolarização (corrente de escuro). Esses canais de Na são cGMP dependentes. Na presença de luz o fotorreceptor reage fotoquímicamente: a rodopsina se descolore e ativa a proteina G, (conhecida como transducina) que por sua vez, estimula a atividade da enzima efetora, uma fosfodiesterase (que que converte o cGMP em GMP). A redução de cGMP inativa os canais de Na causando conrrente de hiperpolarização na membrana do fotorreceptor. Concluimos que o potencial receptor do fotorreceptor é hiperpolarizante e, como consequência, deve reduzir a lberaçao de NT em seu terminal. A luz, causa ainda simultaneamente, a redução de Ca intracelular, fato que estimula a guanili-ciclase, enzima que sintetiza o cGMP antecipando-se para nova situação de ausência de luz.
Faça a comparação entre os mecanismos de açao do fotopigmento e do NT acoplados a proteina G e quando ela é sensibilizada, ocorre inibição na produção do segundo mensageiro. O fato da reaçao fotoquimica utilizar 2º mensageiros significa que é um sistema ampificado de sinal: a sensibildade é tanta que um único foton (unidade de energia luminosa) pode evocar a resposta elétrica no bastonete. Disso podemos deduzir que os bastonetes estão adaptados para a visão em ambientes com baixa iluminação (visão noturna ou escotópica) e que o potencial receptor do fotorreceptor é uma resposta de hiperpolarização (corrente de claro).
aça a comparação entre os mecanismos de açao do fotopigmento e do NT acoplados a proteina G e quando ela é sensibilizada, ocorre inibição na produção do segundo mensageiro. O fato da reaçao fotoquimica utilizar 2º mensageiros significa que é um sistema ampificado de sinal: a sensibildade é tanta que um único foton (unidade de energia luminosa) pode evocar a resposta elétrica no bastonete. Disso podemos deduzir que os bastonetes estão adaptados para a visão em ambientes com baixa iluminação (visão noturna ou escotópica) e que o potencial receptor do fotorreceptor é uma resposta de hiperpolarização (corrente de claro).
Mas como as celulas ganglionares são estimuladas? Veja o esquema a seguir do circuito da retina: os fotorreceptores e as acelulas bipolares não geram potenciais de açao, só as celulas ganglionares. Quando o fotorreceptor gera potencial receptor hiperpolarizante, ocorre redução na liberaçao dos NT. Como consequência, as células bipolares aumentam o nivel de despolarização e excitam as celulas ganglionares a dispararem mais PA pelas suas fibras aferentes (nervo II). Assim as células ganglionares
VIAS VISUAIS
A via visual começa na retina com os fotorreceptores. Os neurônios ganglionares são de 3a ordem e ganham mielina ao deixarem a túnica mais externa para formar o nervo óptico (II par de nervos cranianos). Os dois nervos ópticos se encontram no quiasma óptico e logo se destacam para formar os tratos ópticos e esses terminam no tálamo, nos núcleos geniculados laterais (LGN). Os neurônios talâmicos (4a ordem) partem para o córtex visual através da radiação óptica e atingem os lábios da fissura calcarina no lobo occipital (neurônios de 5a ordem). O córtex visual primário é denominado córtex estriado e funcionalmente está organizada funcionalmente em módulos contendo cada um 150.000 neurônios. No total existiria 2500 módulos sendo que cada um recebe um analisa um determinado aspecto da uma região diminuta do campo visual e que são associadas em outras áreas corticais.
Ao nível do quiasma, ocorre decussação parcial das fibras: as fibras nasais de um mesmo olho cruzam para o lado oposto; já as fibras temporais que se mantêm homolateralmente.
Como conseqüência, no LGN e no córtex de um lado (direito, pe), chegam impulsos da retina nasal do olho esquerdo e da retina temporal do olho direito. Na prática significa que o córtex do lado direito toma consciência do hemicampo visual temporal do olho direito e do hemicampo temporal do olho esquerdo.
As informações da retina têm vários destinos:
As informações originadas na retina não só evocam a percepção visual como também propiciam outras atividades como respostas motoras reflexas e os ritmos biológicos.
1) Fibras retino-geniculadas
São as mais importantes, pois somente estas estão relacionadas com a percepção consciente da visão. Os neurônios talâmicos (de 4a.ordem) projetam-se para o córtex visual localizado no lobo occipital nos lábios da fissura calcarina através da radiação óptica.
2) Fibras retino-hipotalâmicas
Destaca-se do quiasma e ganham o núcleo supraquiasmático do hipotálamo e estão associados com a regulação dos ritmos biológicos;
3) Fibras retino tectais
Projetam-se para os núcleos dos colículos superiores e estão associados com reflexos dos movimentos oculares, reflexo fotomotor direto e consensual.
Quando queremos manter o olhar fixado num determinado objeto, os nervos motores do III, IV e V pares são recrutados para controlar os músculos extrínsecos dos olhos. Entretanto, estamos sujeitos a movimentos da cabeça e do corpo constantemente, tendendo a desfocalizar o objeto de interesse sobre a fóvea. Os órgãos do sentido de equilíbrio (a ser estudado mais adiante) enviam constantemente informações para estes núcleos que corrigem os movimentos dos olhos. Imagine-se sobre o cavalo trotando: sua cabeça estará se movimentando para cima e para baixo. Para evitar a desfocalização de um objeto a sua frente, todas as vezes que a cabeça se abaixar, os olhos se movimentam para cima e vice-versa.
As fibras que chegam na área pré-tectal chegam até o núcleo de Edinger-Westphal e pelo III par de nervos cranianos os neurônios pré-ganglionares chegam até o gânglio ciliar. Os neurônios pós-ganglionares parassimpáticos inervam os músculos esfíncter da pupila, causando a sua contração (miose). A lesão do culo motor causa abolição deste reflexo. Como ocorre o cruzamento das fibras no quiasma, a iluminação do olho de um lado causa, o reflexo fotomotor do olho; daí o nome reflexo consensual, em oposição ao anterior, reflexo fotomotor direto.
Reflexo de Piscar:
O piscar é desencadeado por outros estímulos mas as informações da retina são enviadas também para o núcleo do nervo facial (VII), cujos neurônios motores inervam os músculos orbiculares que causam o fechamento da pálpebra, especialmente quando ocorre aumento de intensidade luminosa. Caso a estimulação seja muito intensa, são enviados impulsos para neurônios motores medulares (Trato teto-espinhal) causando a proteção dos olhos com as mãos.
O campo visual de um olho com a cabeça parada corresponde a 150o; veja-o com um olho fechado. Com os dois olhos abertos, o campo visual fica evidentemente mais ampliado. Fixe um determinado ponto de observação e feche e abra cada olho alternadamente. É fácil perceber que existe uma região central vista por cada olho que se sobrepõe. Este é o campo binocular cuja sobreposição (quando vemos com os dois olhos abertos) nos permite relativizar a profundidade e o relevo dos objetos, proporcionando-nos a visão esteroscópica.




Nenhum comentário:
Postar um comentário